
Лекция № 7
Отдел Покрытосеменные, или Цветковые – Magnoliophyta, или Angiospermae
Цветковые как высший этап эволюции наземных растений. Общая характеристика отдела. Особенности анатомо-морфологического строения. Основные отличия цветка от стробила голосеменных. Цикл воспроизведения. Экология и биология опыления. Двойное оплодотворение.
Основные теории происхождения цветковых. Время, место возникновения и предполагаемые предки.
Разнообразие цветковых растений и их роль в современном растительном покрове. Однодольные и двудольные. Класс Двудольные – Dicotyledones, или Magnoliopsida.
1. Цветковые как высший этап эволюции наземных растений. Общая характеристика отдела. Особенности анатомо-морфологического строения. Основные отличия цветка от стробила голосеменных. Цикл воспроизведения. Экология и биология опыления.
Цветковые – основные вкладчики в биологическую продукцию и биомассу суши (рис.1). Общее число описанных видов – более 250 тыс. видов. Для сравнения голосеменных – около 700 видов, папоротниковидных – около 10 тыс. видов, моховидных – 25 тыс. видов, водорослей – 30 тыс. видов, грибов – около 100 тыс. видов.
В действительности число больше, так как есть виды, неизвестные науке, обитаю во влажных тропических лесах, истребляемых для плантаций кофе, гевеи и бананов.
Рис. 1. Кувшинка белая - вид, занесенный в Красную книгу Бурятии
Цветковые поражают не только морфологическим разнообразием (в «Системе магнолиофитов» А.Л. Тахтаджян указывает 166 порядков, 533 семейства, около 13 тыс. родов), но и разнообразием жизненных форм и широтой спектра условий среды обитания.
У них наиболее развита стратегия патиентности (выносливость к экстремальным условиям среды), и эксплерентности (способность к быстрому заселению незанятых местообитаний).
Размеры цветковых варьируют от гигантских австралийских эвкалиптов (более 100 м) до крохотных (Lemna minor).
Значение цветковых.
В современную эпоху именно покрытосеменные играют главную роль в растительном покрове нашей планеты, занимая огромные площади. Они определяют характер растительности всех континентов. Цветковые являются доминантами, то есть господствуют, во всех типах фитоценозов Земного шара, кроме хвойных лесов, где доминируют голосеменные.
Цветковые растения издавна использовались человеком как пищевые и кормовые, лекарственные, использовались в строительстве и в ремеслах. Многие цветковые сейчас введены в культуру.
Роль цветковых как главных продуцентов биосферы объясняется совершенством организации, кроме перечисленных и особенностями жизненного цикла и еще большей, чем у голосеменных «заботой спорофита о гаметофите». Кратко перечислим важнейшие черты организации цветковых.
Совершенство системы обеспечения водой.
За исключением некоторых родов из Magnoliales и близких к нему порядков (Orimys, Degeneria, Schisandra, Trochodendron и др.), все цветковые имеют развитую проводящую систему с настоящими сосудами. Что позволяет обеспечивать растения влагой даже в условиях ее недостатка в почве. Проводящая система дополняется эффективными покровными тканями (эпидермой и перидермой) и устьичным аппаратом.
Высокая эффективность фотосинтеза.
Развитый листовой аппарат имеет большую поверхность, при достаточном увлажнении располагается многоярусно, образуя 4-5 слоев.
Совершенна внутренняя структура листа. Мезофилл, дифференцированный на столбчатую хлоренхиму и губчатую рыхлую хлоренхиму, пронизан густой сетью проводящих пучков. В зависимости от обеспеченности влагой различаются типы фотосинтеза: С3 у растений, хорошо обеспеченных влагой и С4 у растений сухих местообитаний.
Минеральное питание.
Корневая система разветвленная и может проникать на большую глубину (до 10 м и более). Все цветковые – симбиотрофы и связаны с микоризными грибами, бактериями ризосферы, реже с клубеньковыми азотфиксирующими бактериями (представители семейства Fabaceae).
Генеративные органы.
Совершенство вегетативных органов спорофита сочетается с новым специализированным органом – цветком, обеспечивающим бесполое и половое размножение и образование семян и плодов (рис.2).

Рис. 2. Плоды.
Цветок.
Микроспорофиллы, или тычинки в подавляющем большинстве случаев расчленены на тычиночную нить, связник и пыльники. Однако, у наиболее примитивных Degeneria vitiensis с острова Фиджи он еще листовидной формы. А длинные пыльники, или микроспорангии, располагаются на нижней стороне такой тычинки как у папоротниковидных. Уменьшается число членов андроцея.
Мегаспорофиллы, или плодолистики, у цветковых срослись боковыми краями, образовав полость – гнездо завязи, в котором располагаются семязачатки, а позднее, когда из гинецея образуется плод, и семена (рис.3).
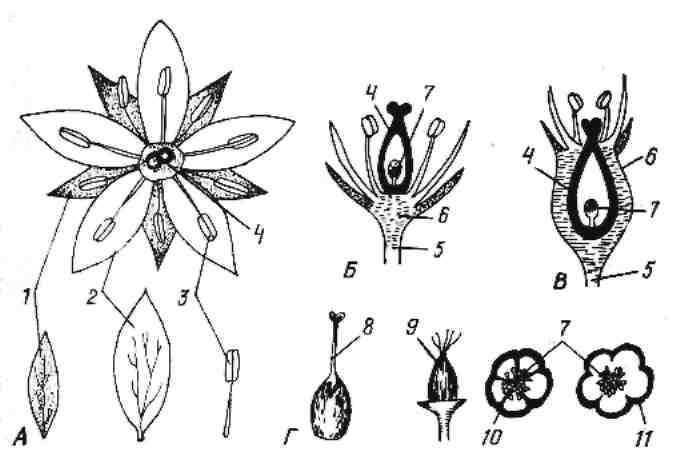
Рис. 3. Строение цветка.
А – цветок (вид сверху): 1 – чашелистики; 2 – лепестки; 3 – андроцей; 4 – гинецей; Б – продольный разрез через цветок (верхняя завязь): 5 – цветоножка; 6 – цветоложе; 7 – семязачаток; В - продольный разрез через цветок (нижняя завязь); Г – гинецей: 8 – столбик; 9 – завязь; 10 – раздельнолепестный венчик; 11 – спайнолепестный венчик.
У Degeneria vitiensis широкие листовидные плодолистики сложены вдоль средней жилки и только соприкасаются в большей части своими краями, но не срастаются. Пестик не имеет разделения на столбик, рыльце и завязь (рис.4).
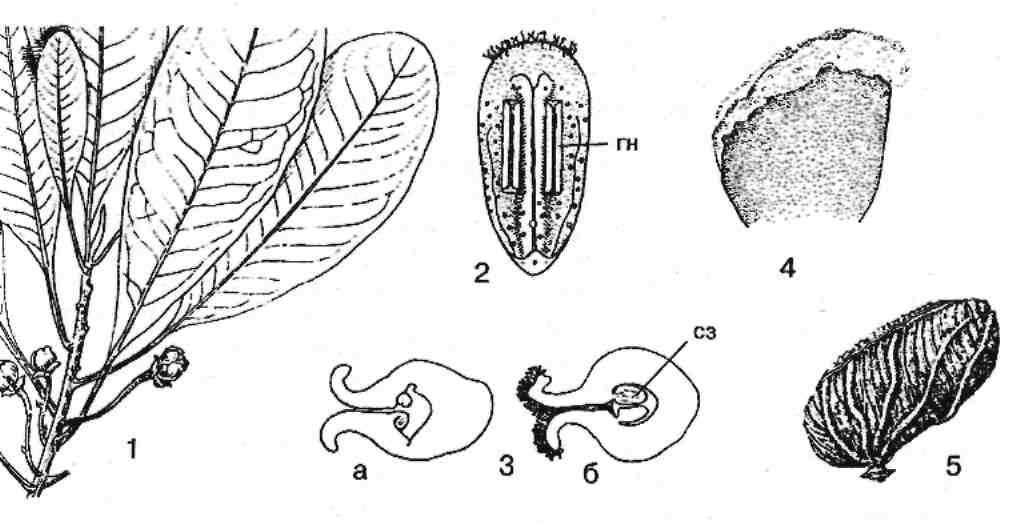
Рис. 4. Дегенерия фиджийская: 1 – цветущая ветвь; 2 – тычинка с нижней стороны (гн – гнезда пыльника); 3 – завязь в поперечном разрезе на ранней (а) и более поздней (б) стадии развития (сз – семязачатки); 4 – пестик; 5 – плод.
Пока плодолистики остаются свободными, гинецей носит название апокарпного. В дальнейшем боковые стенки срастаются, что приводит к синкарпному гинецею. Далее эволюция шла двумя путями:
1. если боковые стенки исчезают, а семязачатки перемещаются на внешние стороны у места их срастания, такой гинецей носит название паракарпного (завязь одногнездная).
2. исчезают боковые стенки, но остается в центре гнезда часть спинных сторон плодолистиков в виде центрального семяносца, на котором располагаются семязачатки, такой гинецей называют лизикарпным.
В строении цветка четко прослеживается две линии специализации: энтомофильная и анемофильная (табл.1).
Таблица 1.
Линии специализации цветковых растений по способу опыления
|
Анемофильная |
Энтомофильная |
|
|
Узкая |
Широкая |
|
|
Увеличивается продукция пыльцы (пыльники крупные) и поверхность рыльца увеличивается (злаки, осоковые). Околоцветник невзрачный или редуцирован. |
Семейства: Бобовые и Орхидные |
Доступна для разных опылителей (сложноцветные): мелкие цветки объединены в соцветия, функционирующие как единая «опылительная система». |
Микроспорогенез и микрогаметогенез.
Микроспорогенез – образование микроспор в микроспорангиях, или пыльцевых мешках, пыльника. Микрогаметогенез – это развитие микроспоры в мужской гаметофит, или пыльцевое зерно.
На первых этапах дифференцировки пыльник состоит из однородной массы клеток и частично обособившейся эпидермы. Затем внутри него выделяются четыре группы фертильных, или спорогенных клеток. Каждая такая группа окружена несколькими слоями стерильных клеток, которые превращаются в стенку пыльцевого мешка. В нее входят и питательные клетки, которые снабжают питательными веществами развивающиеся микроспоры и образуют тапетум, самый внутренний слой стенки. Спорогенные клетки становятся делящимися мейотически диплоидными микроспороцитами, дающими тетраду гаплоидных микроспор. Микроспорогенез завершается образованием одноклеточных микроспор (рис.5).
Во время мейоза образование клеточных стенок либо следует за каждым ядерным делением, либо разделяет протопласты четырех микроспор только после второго деления мейоза. Первый случай типичен для однодольных, второй – для двудольных.
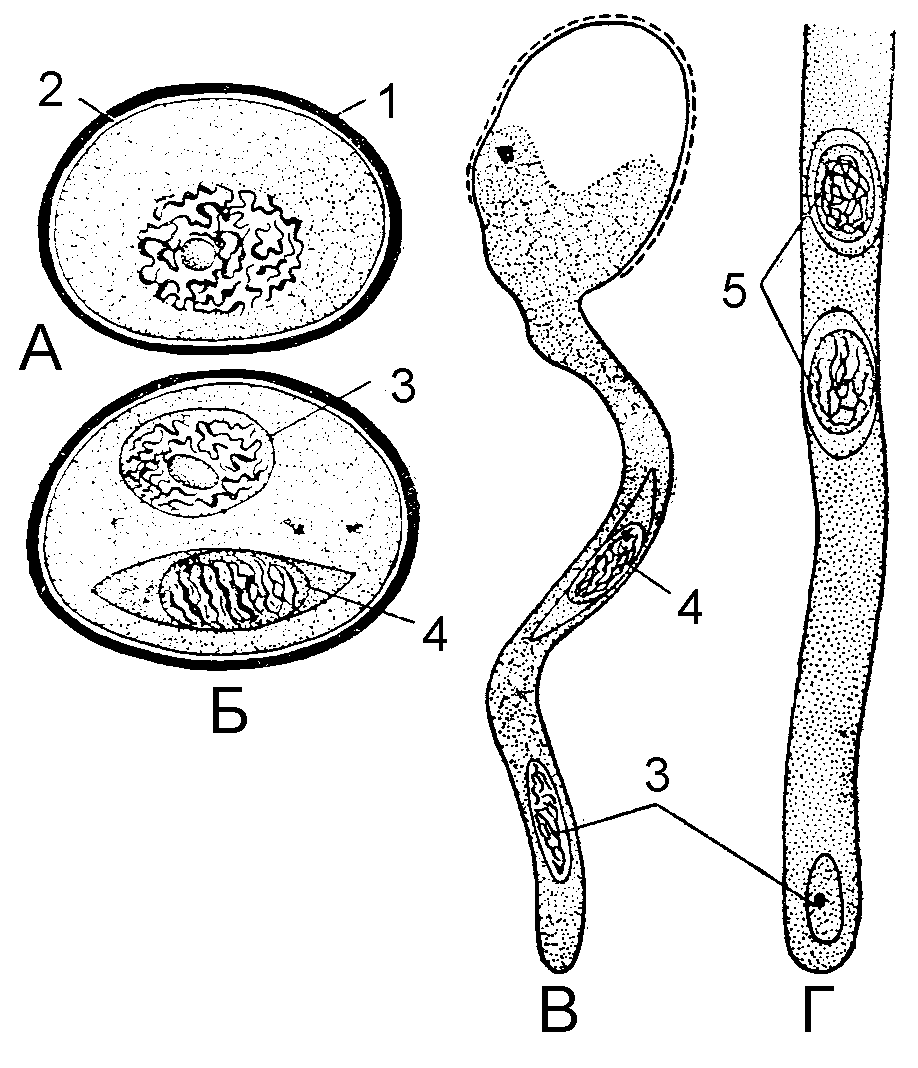
Рис. 5. Микроспора и ее прорастание. А – микроспора; Б – пыльцевое зерно; В – формирование пыльцевой трубки; Г – часть пыльцевой трубки: 1 – экзина; 2 – интина; 3 – вегетативная клетка, 4 – репродуктивная клетка, 5 – спермии.
Следующим этапом является формирование основных признаков пыльцевых зерен. Они окружаются прочной наружной оболочкой – экзиной и целлюлозно-пектиновой внутренней оболочкой – интиной. Экзина состоит из очень стойкого вещества спорополлинина, по-видимому, образуемого частично тапетумом, а частично микроспорой. Интина откладывается протопластом микроспоры. Размеры и форма пыльцевых зерен, как и у спор, сильно варьирует. Их диаметр составляет от менее 20 до более 250 нм. Споры прорастают через шов, проходящий по их центру, а пыльцевые зерна – через поры. Вследствие этого их часто нетрудно различить в ископаемом состоянии.
Микрогаметогенез у покрытосеменных единообразен и начинается с митотического деления одноядерной микроспоры, дающего две клетки внутри ее исходной оболочки. Одна из них называется клеткой трубки, а вторая – генеративной. Мужской гаметофит многих видов ко времени рассеивания пыльцы при вскрывании пыльника находится именно на этой двухклеточной стадии. У других видов генеративное ядро делится, давая начало двум мужским гаметофитам, или спермиям, несколько раньше.
Мегаспорогенез и мегагаметогенез.
Мегаспорогенез – это процесс формирования мегаспоры в нуцеллусе (мегаспорангии). Мегагаметогенез – это развитие мегаспоры в женский гаметофит.
Семяпочка – относительно сложное образование, состоящее из семяножки (фуникулуса), несущей нуцеллус, заключенный в один или два интегумента. В зависимости от вида на плацентах развиваются от одного до многих семязачатков. Развивающаяся семяпочка первоначально состоит целиком из нуцеллуса, но вскоре возникают один или два покровных слоя (интегумента) с небольшим отверстием, микропиле, на одном конце (рис.6).
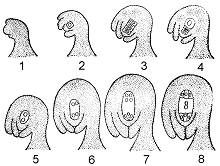
Рис. 6. Схема формирования семязачатка и зародышевого мешка.
1, 2, 3, 4 – развитие нуцеллуса, обособление и мейоз клетки археспория, отмирание трех мегаспор; 5, 6, 7, 8 – развитие из мегаспоры (оставшейся) женского гаметофита – зародышевого мешка.
На ранней стадии развития семязачатка в нуцеллусе возникает единственный диплоидный мегаспороцит. Он делится митотически, давая четыре гаплоидные мегаспоры, обычно расположенные линейной тетрадой. Этим завершается мегаспорогенез. Три мегаспоры обычно разрушаются, а четвертая, наиболее удаленная от микропиле, развивается в женский гаметофит.
Функциональная мегаспора вскоре начинает увеличиваться за счет нуцеллуса, а ее ядро трижды делится митотически. В конце третьего митоза восемь дочерних ядер располагаются по четыре двумя группами – вблизи микропилярного конца мегагаметофита, а также на противоположном, халазальном, конце. По одному ядру из каждой группы мигрирует к центру восьмиядерной клетки; они называются полярными. Три оставшихся у микропилярного конца ядра образуют яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. У халазального конца также идет формирование вокруг расположенных здесь ядер клеточных оболочек, и возникают так называемые клетки-антиподы. Полярные ядра остаются в двуядерной центральной клетке. Такая восьмиядерная семиклеточная структура является зрелым женским гаметофитом, называемым зародышевым мешком.
Опыление и двойное оплодотворение.
При вскрывании (рассеивании содержимого) пыльников пыльцевые зерна переносятся на рыльце многими способами; процесс этого переноса называется опылением.
У цветковых существуют разнообразные приспособления к перекрестному опылению и ограничению самоопыления: двудомность, самонесовместимость (ингибирование прорастания собственной пыльцы), дихогамия – протандрия и протогиния (неодновременное созревание тычинок и пестиков), херкогамия (пространственное разобщение пыльцы и рыльца, как у губоцветных). При этом самоопыление у многих видов возможно как «запасной вариант».
Вступив в контакт с рыльцем, пыльцевые зерна поглощают дополнительное количество воды из клеток его поверхности (по градиенту водного потенциала). После насыщения водой они прорастают в пыльцевую трубку. Генеративная клетка, если она еще не поделилась, вскоре делится, образуя два спермия. Проросшее пыльцевое зерно с ядром трубки и двумя спермиями и соответствует зрелому мужскому гаметофиту (рис.7).
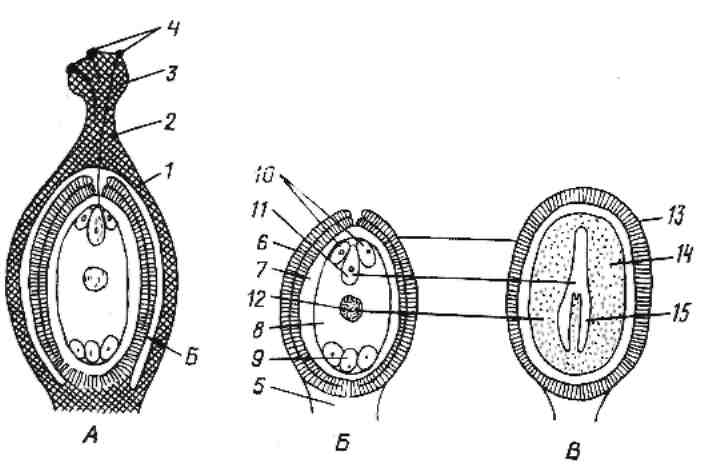
Рис. 7. Оплодотворение (А), строение семяпочки (Б) и семени (В). 1 – завязь, 2 – столбик, 3 – рыльце, 4 – прорастающая пыльца, 5 – семяножка, 6 – покровы семяпочки, 7 – остатки нуцеллуса, 8 – зародышевый мешок, 9 – три антиподы, 10 – две синергиды, 11 – яйцеклетка, 12 – вторичное ядро зародышевого мешка, 13 – семенная кожура, 14 – эндосперм, 15 – зародыш.
Рыльце и столбик структурно-физиологически устроены так, чтобы облегчить прорастание пыльцевого зерна и рост пыльцевой трубки. Поверхность многих рылец образована главным образом железистой тканью (рыльцевая ткань), выделяющей сахаристый раствор. Эта ткань связана с семяпочкой проводниковой (трансмиссионной) тканью, своего рода дорожкой сквозь столбик для растущих пыльцевых трубок. В некоторых столбиках проводниковой тканью выстланы изнутри открытые канальцы, и пыльцевые трубки растут либо поверх этой выстилки, либо среди ее клеток. Однако у большинства покрытосеменных столбики выполненные (сплошные) с одним или несколькими тяжами и проводниковой ткани между рыльцем и семяпочками. Пыльцевые трубки здесь в зависимости от вида растения растут либо среди ее клеток, либо внутри их толстых стенок.
Обычно пыльцевая трубка входит в семяпочку через микропиле и проникает в одну из синергид, которая начинает отмирать вскоре после опыления, но раньше, чем пыльцевая трубка достигает зародышевого мешка.
Два спермия и ядро трубки выпускаются в эту синергиду через открывающуюся в пыльцевой трубке субтерминальную пору. Затем ядро одного из спермиев проникает в яйцеклетку, а другого – в центральную клетку, где сливается в двумя полярными ядрами. В этом главное отличие от голосеменных, у которых функционален только один из двух спермиев, сливающийся с яйцеклеткой, в то время как второй дегенерирует. Слияние одного спермия с яйцеклеткой, а другого с полярными ядрами – двойное оплодотворение – и представляет собой уникальную особенность покрытосеменных. В результате слияния одного спермия с яйцеклеткой образуется диплоидная зигота. Соединение другого спермия с двумя полярными ядрами, то есть тройное слияние, дает триплоидное первичное ядро эндосперма.
Ядро клетки трубки тем временем отмирает, оставшаяся синергида и антиподы отмирают вместе с ним или на ранних стадиях дифференцировки зародышевого мешка.
Следует отметить, что у представителей некоторых семейств (в особенности у сложноцветных) распространено явление апомиксиса – девственного развития семян без оплодотворения.
После двойного оплодотворения начинается несколько процессов: первичное ядро эндосперма делится, образуя эндосперм; зигота развивается в зародыш; интегументы превращаются в семенную кожуру, стенка завязи и связанные с ней структуры формируют плод. Развитие семяпочки в семя сопровождается превращением завязи (а иногда и других частей цветка или соцветия) в плод.
В ходе этого стенка завязи (околоплодник, или перикарпий) часто утолщается и дифференцируется на отдельные слои – внешний экзокарпий (внеплодник), средний мезокарпий (межплодник) и внутренний эндокарпий (внутриплодник) (иногда только на экзо- и эндокарпий), - обычно лучше заметные в мясистых, чем в сухих, плодах.
Жизненный цикл покрытосеменных.
Гаметофиты цветковых очень сильно редуцированы в размерах – больше, чем у любых других разноспоровых растений, включая голосеменные. Мужской в зрелом состоянии состоит только из трех клеток; а женский, погруженный в течение всей жизни в ткани спорофита, у большинства видов только из семи. Антеридии и архегонии отсутствуют. Опыление непрямое, т.е. пыльца откладывается на рыльце, а затем пыльцевая трубка доставляет два неспособных самостоятельно двигаться спермия к женскому гаметофиту. После оплодотворения семяпочка дает семя, заключенное в завязи, которая одновременно (иногда вместе с сопутствующими ей структурами) развивается в плод.
Семена цветковых способны к длительному покою и имеют разнообразные приспособления к распространению. Многие виды способны размножаться вегетативно (корневищами, клубнями, луковицами и пр.).
Все эти качества цветковых и объясняют их широкое распространение, хотя в некоторых районах они уступают голосеменным. Однако при интенсивном влиянии человека голосеменные не выдерживают конкуренции и уступают более пластичным цветковым.
2. Основные теории происхождения цветковых. Время, место возникновения и предполагаемые предки.
По выражению Тахтаджяна, гипотезы - это строительные леса для здания теоретического знания. Хотя, к сожалению, далеко не в любых «лесах» это «здание» может быть построено. И видимо происхождению цветковых так и суждено остаться «отвратительной тайной», выраженное Ч. Дарвином в письме Дж. Гукеру в 1879 г.
Во времена Дарвина о происхождении цветковых было известно очень мало. Изучение палеоботанической летописи только начинались. На основании этих скудных сведений Дарвин считал, что цветковые появились внезапно в меловом периоде и сразу во всем разнообразии.
К концу 40-х гг. XX в., когда сформулировал свою гипотезу происхождения цветковых А.Л. Тахтаджян, палеоботанических сведений было еще недостаточно. Толчок к разработке гипотезы дали экспедиции Тахтаджяна в юго-восточные районы Китая, где он обнаружил роды магнолиевых с гомоксильной («трахеидной») древесиной, свойственной голосеменным. На палеографической карте мезозоя территория юго-восточного Китая называлась Катазией и гипотеза Тахтаджяна была названа катазионной. Все положения своей гипотезы Тахтаджян вывел дедуктивно, как плод «игры воображения» (табл. 2).
Спустя 30 лет, когда в 70-е гг. Родилась гипотеза С.В. Мейена, палеоботаническая летопись уже была пополнена множеством данных. Были обнаружены первые ископаемые цветковые растения, которые были непохожи на магнолиевые с гомоксильной древесиной. Эти первые цветковые были распространены по всему тропическому поясу, откуда они расселились в высокие широты к северу и к югу. Гипотеза Мейена получила название гипотезы «тропической помпы» (фитоспрединга). Таким образом, если Тахтаджян основывался на данных о современных закономерностях распределения цветковых - «живых ископаемых», то Мейен - на достаточно большой палеоботанической информации.
Кроме того, во времена формулирования гипотезы Тахтаджяна дарвинизм и синтетическая теория эволюции (СТЭ) переживали апогей развития (особенно в СССР). К началу 70-х гг. популярность дарвинизма резко упала. Стало очевидным, что СТЭ не в состоянии объяснить сформировавшееся разнообразие видов растений и животных, так как модель «случайные ненаправленные мутации и отбор полезных изменений условиями среды» описывает слишком медленный процесс. В это время получают развитие ранее отвергавшиеся не дарвинистские варианты объяснения эволюции - номогенез (развитие по программе, заложенной в самой природе живого вещества) и сальтационизм (допущение крупных скачкообразных изменений форм). Этот новый уровень развития теории эволюции отражен в гипотезе Мейена (табл. 2).
Таблица 2
Основные антиномии гипотез происхождения цветковых.
|
Вопрос |
Гипотеза |
|
|
|
катазионная |
фитоспрединга |
|
Время происхождения |
нижний и др. мел |
нижний-верхний мел |
|
Время расселения |
верхний мел |
нижний-верхний мел |
|
Место происхождения |
Катазия (Юго-Восточная Азия) |
тропический пояс |
|
Экологические условия |
горы |
различные, преимущественно равнинные |
|
Предки |
семенные папоротники |
беннеттиты |
|
Древние формы |
вечнозеленые деревья с одиночными крупными цветками типа магнолиевых |
различные древесинные растения с невзрачными цветками, собранными в соцветия |
|
Способ опыления древних форм |
энтомофилия и особенно кантарофилия (жуками) |
факультативное: энтомофилия, анемофилия, самоопыление |
|
Основной эволюционный механизм возникновения разнообразия |
Синтетическая теория эволюции. Адаптации к различным горным местообитаниям (по высоте над уровнем моря, экспозиции). Сочетание мутаций и направленно отбора. Популяционные волны. Генетический дрейф. Чередование периодов изоляции локальных популяций (демов) и гибридизации. |
Сальтации: выживание организмов с крупными изменениями формы неадаптивного характера, откачивание их «тропической помпой» в высокие широты и адаптационные изменения в соответствии с СТЭ. |
Уязвимым моментом обеих гипотез является представления о предках цветковых. Тахтаджян считает таковыми семенные папоротники, но выводит это благодаря жестким дедукциям, путем последовательного исключения всех прочих групп голосеменных. Естественно, что невозможно исключить или привлечь те группы, которые еще неизвестны, а такое вполне вероятно.
Мейен считает предками цветковых беннеттитовые, стробил которых был обоеполым. При этом несоответствие строения макроспорофиллов беннеттитовых и цветковых он пытается преодолеть гипотезой о «переносе фруктификаций», т.е. о достаточно сложных миграциях генеративных органов внутри структуры «процветка». В.Н. Тихомиров (1987) назвал этот элемент гипотезы «экстравагантным».
Быстрое развитие покрытосеменных растений, возможно, было вызвано резкими изменениями климатических условий на Земле: освещенности и влажности воздуха в середине мелового периода. Чрезвычайная эволюционная пластичность, способность к адаптации, надежная защита семяпочек и семени обусловили быстрое распространение покрытосеменных и вытеснение ранее господствовавших групп высших споровых растений.
Первые покрытосеменные были, скорее всего, древесными растениями с преобладанием моноподиального ветвления. Позже в разных линиях эволюции появились кустарниковые и травянистые жизненные формы, вначале многолетние, а затем однолетние. Выработалось огромное разнообразие вегетативных органов, особенно листьев, появились многочисленные метаморфозы, а также бесконечно разнообразные в своем строении и окраске цветки и плоды. Основными органами цветка являются: гинецей, андроцей, околоцветник, расположенные на цветоложе.
Покрытосеменные по сравнению с голосеменными имеют еще более редуцированные женский (зародышевый 8-ядерный мешок) и мужской (пыльца) заростки; при развитии мужского гаметофита никогда не образуются проталлиальные клетки (остаток вегетативного тела заростка).
Цветок представляет собой целостную систему. Для примитивного цветка было характерно наличие более менее замкнутых вместилищ, образуемых плодолистиками, в полости которых развивались семязачатки. Многие типы цветков современных цветковых имеют примитивные черты организации и принимаются за первичные - типа магнолиевых с удлиненным цветоложем и развитым околоцветником, беспокровные цветки типа троходендровых или цветок со стерильными частями между тычинками и пестиками, как у Eupomatia.
Эволюция цветка, на строении которого и базируется главным образом систематика покрытосеменных, шла от первичных актиноморфных цветков со спиральным расположением неопределенных в числе органов цветка с верхней завязью и однопокровными семяпочками - к цветкам циклическим, с вполне определенным числом членов, часто более менее сросшихся, к цветкам зигоморфным с нижней завязью, часто с редукцией числа семяпочек. Эти изменения происходили в разных эволюционных рядах с различной скоростью и последовательностью.
Такова современная точка зрения на происхождение цветковых.
Однако эти представления сложились далеко не сразу. На рубеже XIX-XX вв. главенствовала так называемая псевдантиевая теория, принадлежащая двум известным ботаникам - Р. Веттштейну и А. Энглеру. Суть этой теории в том, что цветковые связывались своим происхождением с хвойными растениями, ветроопыляемыми и имеющими раздельнополые шишки (пыльниковые и семенные). Поэтому наиболее примитивными цветковыми считались ветроопыляемые с редуцированным околоцветником и почти всегда с раздельнополыми цветками, так называемые сережкоцветные (березовые, буковые, крапивные, коноплевые, тутовые и др.). Из голосеменных наиболее близкими к цветковым полагались оболочкосеменные, как бы переходная группа. Их признаки: остатки былой обоеполости, наличие покровов у стробилов, похожего на околоцветник, наличие сосудов. А наиболее близким к голосеменным из цветковых считалось семейство казуариновые, у которых мужские и женские цветки собраны в отдельные соцветия, напоминающие шишку, причем мужской цветок состоит из одной тычинки и двух прицветничков (но лишен околоцветника), а женский из двух сросшихся плодолистиков, тоже с 2 прицветничками. По мнению авторов теории, раздельнополые соцветия объединились и образовали обоеполый цветок, по существу, являющийся целым соцветием. Эта теория получила название псевдантиевая.
Опыление насекомыми, считали авторы теории, пришло позднее и вызывало появление околоцветника.
Но уже у начале XX в. два систематика, Г. Галлир и Ч. Бесси, независимо друг от друга пришли к диаметрально противоположным выводам. Они справедливо полагали, что первичные цветковые были насекомоопыляемыми, имели обоеполый цветок (стробил) с ярким околоцветником, а так называемые сережкоцветные отнюдь не являются наиболее древними, первичными. Эта теория рассматривала цветок как обоеполый стробил, а не как соединение двух раздельнополых стробил. Теория получила название эвантиевой.
Но ни Галлир, ни Бесси не были палеоботаниками и не привлекали к своей теории данные о вымерших растениях, поэтому, когда вслед за их работами появилось исследование Н. Арбера и Д. Паркина, изучавших обоеполые стробилы беннеттитов, теория Галлира и Бесси получила неоспоримое подтверждение в виде ископаемого материала. Арбер и Паркин назвали свою теорию стробилярной и выводили цветковые из беннеттитов.
Начиная с 30-х гг. XX в. получила распространение теломная теория, отвергающая плодолистик как орган листовой природы. Предложены многочисленные варианты формирования разных типов цветков из различных предковых структур, сводимых к исходной системе стерильных и фертильных теломов.
В системах, принятых в настоящее время, в начале системы ставятся семейства с хорошо развитыми многочисленными, раздельнолепестными, двупокровными, энтомофильными цветками, как, например, семейства магнолиевые, лютиковые, кувшинковые, барбарисовые и другие, так называемые многоплодниковые. Семейства с однопокровными и беспокровными цветками считаются вторично упрощенными, так как многие имеют ценокарпный гинецей, нижнюю завязь - признаки не примитивные. Таковы системы ботаников Буша, Гроссгейма, Козо-Полянского, Кузнецова, Тахтаджяна, Галлира, Пула, Шафнера, Гетчинсона, Шоо.
3. Разнообразие цветковых растений и их роль в современном растительном покрове. Однодольные и двудольные. Класс Двудольные – Dicotyledones, или Magnoliopsida.
Отдел Покрытосеменные делится на два класса: Двудольные (Dicotyledones, или Magnoliopsida) и Однодольные (Monocotyledones, или Liliopsida). В большинстве современных систем принимается, что однодольные произошли от примитивных первичных двудольных.
По последней системе А.Л. Тахтаджяна эти классы подразделяются на 11 подклассов, 8 из которых принадлежат двудольным, 3 - однодольным. Подклассы включают порядки, состоящие из семейств.
4. Класс Двудольные – Dicotyledones, или Magnoliopsida.
Хотя наличие 2 семядолей в семени и является характерным признаком, он и является характерным признаком, но не является абсолютным. Например, из семейства лютиковые у некоторых представителей 1 семядоля, у очень древних - 3-4.
Листья с перистым или пальчатым жилкованием. Простые и сложные, расчлененные на пластинку и черешок, нередко с прилистниками. Листовых следов в стебле обычно 1-3.
Проводящая система стебля имеет кольцевое строение, а проводящая пучки открытого типа (исключение – семейство нимфейные); в стебле хорошо выражена кора и сердцевина.
При прорастании семени зародышевый корешок развивается в главный корень, от которого отходят боковые. Таким образом, корневая система обычно стержневая, но у ряда травянистых форм из числа примитивных семейств (лютиковые) может быть мочковатой.
Цветки пяти-четырехчленные, реже трехчленные. Обычно (но не всегда) с двойным околоцветником.
Среди жизненных форм у двудольных наблюдается большое разнообразие, причем древесные формы обычно (но не всегда) первичные, а травянистые возникли из них, хотя есть отдельные исключения, когда наоборот, травянистые дали начало вторично древовидным формам.
Класс двудольных в настоящее время подразделяется на 8 подклассов, объединяющих 190 тыс. видов растений.
1. Подкласс Magnoliidae
Представлен в основном древесными растениями, реже травами, водными растениями или паразитами. Для анатомии проводящей системы характерно либо отсутствие сосудов, либо наличие примитивных сосудов с лестничной перфорацией; нередки секреторные клетки, выделяющие эфирные масла, смолы, бальзамы. Цветки обоеполые, спиральные, или спироциклические, с неопределенным андроцеем и большей частью апокарпным гинецеем; семена с эндоспермом, иногда с периспермом и маленьким зародышем. Подавляющее большинство представителей – тропические и субтропические растения. Порядки: Магнолиецветные, Лавроцветные, Бадьяноцветные, Нимфейноцветные, Лотосоцветные.
2. Подкласс Ranunculidae
Близкие к Магнолиидам по ряду признаков, но более высокоорганизованные. Среди них преобладают травы, уже имеющие сосуды, а секреторные клетки становятся редкостью. Цветки тоже обычно обоеполые, спиральные, или спироциклические, андроцей большей частью многочисленный. Как и гинецей, последний обычно апокарпный. Семена с эндоспермом и маленьким зародышем. Порядки: Лютикоцветные, Макоцветные.
3. Подкласс Caryophyllidae
Большинство составляют травянистые растения, полукустарники, кустарники и изредка некрупные древесные формы. Листья простые, цельные. Для проводящей системы характерно наличие сосудов с простой перфорацией. Цветки обоеполые, актиноморфные, изредка раздельнополые или безлепестные, гинецей ценокарпный и лишь изредка апокарпный; семена с согнутым зародышем, окруженным периспермом.
4. Подкласс Dilleniidae
Относятся деревья, кустарники и травы с листьями цельными или по-разному расчлененными. Для проводящей системы характерно наличие сосудов то с лестничной, то с простой перфорацией. Цветки обоеполые и раздельнополые, с двойным околоцветником или иногда безлепестные, у более примитивных семейств спироциклические; андроцей нередко многочисленный, гинецей у примитивных форм апокарпный, у более продвинутых ценокарпный, семена с эндоспермом. Порядки: Чайноцветные, Фиалкоцветные, Мальвовые, Вересковые, Первоцветные, Ивоцветные, Капероцветные, Тыквенные, Крапивные.
5. Подкласс Rosidae
Относятся деревья, кустарники и травы с листьями цельными, расчлененными, сложными. Для анатомии проводящей системы характерны сосуды с простой перфорацией и реже с лестничной. Цветки обоеполые, чаще с двойным околоцветником или реже безлепестные, актиноморфные и зигоморфные; андроцей от многочисленного до определенного, гинецей апокарпный у более примитивных семейств и ценокарпный у более продвинутых; семена с эндоспермом или уже без него. Порядки: Розоцветные, Бобовоцветные, Кизиловые, Аралиевые, Ворсянковые.
6. Подкласс Lamiidae
Жизненная форма – травы, реже деревья или кустарники с листьями либо цельными, либо разнообразно расчлененными. Для анатомии проводящей системы характерны сосуды чаще с простой перфорацией, реже с лестничной. Цветки чаще всего обоеполые, сростнолепестные и нередко зигоморфные, андроцей в определенном числе, нередко меньшим, чем число частей околоцветника, гинецей всегда ценокарпный (паракарпный), семена нередко без эндосперма. Порядки: Горечавковые, Синюховые, Пасленовые, Бурачниковые, Норичниковые, Губоцветные.
7. Подкласс Asteridae
Принадлежат многочисленные травы, реже полукустарники, кустарники, деревья. Цветки чаще всего обоеполые, реже однополые, сростнолепестные, иногда зигоморфные, с андроцеем в определенном числе, обычно равном лепесткам, гинецей всегда ценокарпный, завязь нижняя.
Все перечисленные подклассы представляют собой насекомоопыляемую линию эволюции двудольных. Хотя у отдельных двудольных ветроопыляемых растений произошли некоторые изменения в строении цветков с потерей околоцветника, образованием большого количества сухой легкой пыльцы, выработкой соцветий, в классе двудольных есть особая ветвь, где ветроопыление начинает преобладать над насекомоопылением. Эти порядки и семейства объединяются в подкласс Гамамелидид.
8. Подкласс Hamamelididae.
Относятся главным образом древесные растения или кустарники, травы значительно реже. Для проводящей системы характерны сосуды с лестничной или простой перфорацией. Исключение – древний порядок Trochodendrales, нет сосудов, есть трахеиды. Цветки Гамамелидид в большинстве случаев анемофильные, мелкие, с невзрачным околоцветником или без него, большей частью раздельнополые, с многочисленным неопределенным андроцеем у более примитивных представителей и более определенным у продвинутых, гинецей ценокарпный, плоды обычно односемянные, семена с эндоспермом или без него. Порядки: Троходендроцветные, Гамамелидоцветные, Букоцветные, Березоцветные, Орехоцветные.